ņä£ ļĪĀ
ļ░£ņĢö Ļ│╝ņĀĢņŚÉ ņ׳ņ¢┤ ņĀĢņāü ņäĖĒżĻ░Ć ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ņāØļ¼╝ĒĢÖņĀüņØĖ ļŖźļĀźļōżņØä Ļ░¢Ļ▓ī ļÉśĻ│Ā, ņØ┤ļ¤¼ĒĢ£ ņĢöņäĖĒżņØś ĒŖ╣ņä▒ļōż(hallmarks of cancer)ņØĆ ņĢöņØś ņóģļźśļź╝ ļ¦ēļĪĀĒĢśĻ│Ā Ļ│ĄĒåĄņĀüņ£╝ļĪ£ ļ│┤ņØ┤ļŖö Ēśäņāüņ£╝ļĪ£ Ļ░äņŻ╝ļÉ£ļŗż[1]. ĒŖ╣Ē׳, ņäĖĒż ņŚÉļäłņ¦Ć ĒÖ£ņÜ®ņØś ļ│ĆĒÖö Ēś╣ņØĆ ņŚÉļäłņ¦ĆļīĆņé¼ņØś ņ×¼ĒÄĖņä▒ņØĆ ņĄ£ĻĘ╝ņŚÉ ņČöĻ░ĆļÉ£ ņĢöņØś ņżæņÜö ĒŖ╣ņä▒ņ£╝ļĪ£ņä£, ņĢöņØ┤ ĻĘĖ ņóģļźśņÖĆ ņøÉņØĖņØ┤ ļŗżņ¢æĒĢśĻ▓ī ņĪ┤ņ×¼ĒĢ©ņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā, ņØ┤ļ¤¼ĒĢ£ ņä▒Ē¢źņØĆ ļ¬©ļōĀ ņĢöņŚÉņä£ Ļ│Ąņ£ĀļÉ£ļŗż[2]. ņĢöņäĖĒż ļīĆņé¼ņØś ĒŖ╣ņ¦ĢņĀüņØĖ ļ│ĆĒÖöļĪ£ ļīĆĒæ£ņĀüņ£╝ļĪ£ Ļ▒░ļĪĀļÉśļŖö Ļ▓āņØ┤ ņÖĆļ▓äĻĘĖ ĒÜ©Ļ│╝(Warburg effect)ņØ┤ļŗż. ņØ┤Ļ▓āņØĆ ņĢöņäĖĒżĻ░Ć ņé░ņåīĻ░Ć ņČ®ļČäĒ׳ ņĪ┤ņ×¼ĒĢśļŖö ņĪ░Ļ▒┤ņ×äņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā ņŚÉļäłņ¦Ć ļīĆņé¼ņĀüņ£╝ļĪ£ ĒÜ©ņ£©ņØ┤ ļåÆņØĆ ņé░ĒÖöņĀü ņØĖņé░ĒÖö(oxidative phosphorylation, OXPHOS) Ļ│╝ņĀĢļ│┤ļŗż ĒĢ┤ļŗ╣ Ļ│╝ņĀĢ(glycolysis)ņØä ņäĀĒśĖĒĢśļŖö ĒśäņāüņØä ņØ╝ņ╗½ļŖö Ļ▓āņ£╝ļĪ£ņä£ ĒśĖĻĖ░ņä▒ ĒĢ┤ļŗ╣ ņ×æņÜ®(aerobic glycolysis)ņ£╝ļĪ£ļÅä ļČłļ”░ļŗż[3]. OXPHOS Ļ│╝ņĀĢņØä ĒåĄĒĢ┤ ļ¦ÄņØĆ ņ¢æņØś adenosine triphosphate(ATP)(ĒżļÅäļŗ╣ ĒĢ£ ļČäņ×Éļŗ╣ 36Ļ░£ņØś ATP)ļź╝ ņāØņé░ĒĢśļŖö ļīĆņŗĀ ņāüļīĆņĀüņ£╝ļĪ£ ņŚÉļäłņ¦Ć ĒÜ©ņ£©ņØ┤ ļ¢©ņ¢┤ņ¦ĆļŖö ĒĢ┤ļŗ╣ Ļ│╝ņĀĢ(ĒżļÅäļŗ╣ ĒĢ£ ļČäņ×Éļŗ╣ 2Ļ░£ņØś ATP)ņØä ņ▒äĒāØĒĢ©ņ£╝ļĪ£ņŹ© ņĢöņäĖĒżļŖö ļ│┤ļŗż ļ╣Āļź┤Ļ▓ī ATPļź╝ ņāØņé░ĒĢśĻ│Ā, ĒÖ£ņä▒ ņé░ņåīņóģ(reactive oxygen species, ROS)ņØś ļ░£ņāØņØä ņżäņØ┤Ļ│Ā, ņĀĆņé░ņåīņØś ņóģņ¢æļ»ĖņäĖĒÖśĻ▓Į(tumor microenvironment)ņŚÉņä£ ļŗ╣ļīĆņé¼ņØś ņĢłņĀĢņä▒ņØä Ļ░¢ņČöĻ│Ā, ņżæņÜöĒĢśĻ▓īļŖö ļŗ╣ņØś ņżæĻ░äļīĆņé¼ņ▓┤Ļ░Ć ņŚ¼ļ¤¼ ņāØĒĢ®ņä▒ Ļ▓ĮļĪ£ļĪ£ ņ£Āņ×ģļÉśļŖö Ļ▓āņØä ļŖśļĀż, ĻČüĻĘ╣ņĀüņ£╝ļĪ£ ņĢöņäĖĒż ņä▒ņןņŚÉ ĒĢäņÜöĒĢ£ ļē┤Ēü┤ļĀłņśżĒŗ░ļō£(nucleotide), ņĢäļ»ĖļģĖņé░, ņ¦Ćļ░® ļō▒ņØś ļ░öņØ┤ņśżļ¦żņŖż(biomass) ĒĢ®ņä▒ņØä ĒåĄĒĢ£ ļÅÖĒÖöņ×æņÜ®(anabolism)ņŚÉ ĻĖ░ņŚ¼ĒĢśĻ▓ī ļÉśļŖö ņāØļ¼╝ĒĢÖņĀüņØĖ ņןņĀÉņØä Ļ░Ćņ¦ĆĻ▓ī ļÉ£ļŗż.
ņĢĮ 100ņŚ¼ ļģä ņĀä Warburg ļō▒[4]ņŚÉ ņØśĒĢ┤ ļ│Ė ņØ┤ļĪĀņØ┤ ņ▓śņØī ņĀ£ĻĖ░ļÉĀ ļŗ╣ņŗ£ņŚÉļŖö, ņĢöņäĖĒż ļé┤ņŚÉņä£ļŖö ļ»ĖĒåĀņĮśļō£ļ”¼ņĢäņØś ĻĖ░ļŖź ļČĆņĀäņ£╝ļĪ£ ņØĖĒĢ┤ OXPHOS Ļ│╝ņĀĢņØ┤ ņåÉņāüļÉśĻ▒░ļéś ņ¢ĄņĀ£ļÉśĻ│Ā ņ׳ļŗżĻ│Ā ņāØĻ░üņØä ĒĢśņśĆļŗż. ĻĘĖļ¤¼ļéś ņØ┤ĒøäņŚÉ ņĢöņäĖĒż ļé┤ņŚÉņä£ļÅä ņĀĢņāü ņäĖĒżņÖĆ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ OXPHOS Ļ│╝ņĀĢņØ┤ ņØ╝ņ¢┤ļéĀ ņłś ņ׳ļŗżĻ│Ā ņĢīĻ▓ī ļÉśņŚłņ£╝ļ®░[5-8], ņĢöņäĖĒżĻ░Ć ļåōņØĖ ļŗżņ¢æĒĢ£ ĒÖśĻ▓ĮņŚÉ ļö░ļØ╝ ĒĢ┤ļŗ╣ ņ×æņÜ®Ļ│╝ OXPHOS ņ×æņÜ® Ļ░äņŚÉ ļīĆņé¼ ņĀäĒÖś(metabolic switch)ņØä ĒåĄĒĢ┤ ļŖźļÅÖņĀüņ£╝ļĪ£ ļīĆņ▓śĒĢĀ ņłś ņ׳ņØīņØä ņĢīĻ▓ī ļÉśņŚłļŗż. ņĢöņäĖĒż ļé┤ņŚÉņä£ļŖö ļŗ╣ ĒØĪņłś ņ”ØĻ░Ćļź╝ ĒåĄĒĢ┤ ļŗ╣ļīĆņé¼Ļ░Ć ĒÖ£ņä▒ĒÖöļÉśņ¢┤ ņ׳ņ¦Ćļ¦ī, ļŗ╣ ņØ┤ņÖĖņØś ļŗżļźĖ ņśüņ¢æļČä ĒØĪņłśļź╝ ĒåĄĒĢ┤ ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ tricarboxylic acid(TCA) ĒÜīļĪ£Ļ░Ć ĒÖ£ņä▒ĒÖöļÉśĻĖ░ļÅä ĒĢ£ļŗż.
Ļ│╝Ļ▒░ņŚÉļŖö ņĢöņāØļ¼╝ĒĢÖņŚÉ ņ׳ņ¢┤ ņĢöļīĆņé¼(cancer metabolism)Ļ░Ć ņŻ╝ļźś ĒĢÖļ¼Ėņ£╝ļĪ£ņä£ Ēü░ Ļ┤Ćņŗ¼ņØä ļ░øņ¦Ć ļ¬╗ĒĢśļŗżĻ░Ć, ņĄ£ĻĘ╝ņŚÉ ļōżņ¢┤ņä£ ņĢöļīĆņé¼Ļ░Ć ņóģņ¢æņŗĀĒśĖņĀäļŗ¼(oncogenic signal transduction)ņØ┤ļéś ņóģņ¢æļ®┤ņŚŁ(tumor immunity)Ļ│╝ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ ņĢöņāØļ¼╝ĒĢÖņØś ĒĢäņłś ņÜöņåīļĪ£ ĻĘĖ Ļ┤Ćņŗ¼ņØ┤ ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ļŗż. ĒŖ╣Ē׳ļéś ņĢöļīĆņé¼ ņżæņŚÉņä£ļÅä ņ¦Ćļ░®ņé░ ļīĆņé¼ņÖĆ Ļ┤ĆļĀ©ĒĢśņŚ¼ ĻĘĖ ņŚŁĒĢĀņŚÉ ļīĆĒĢ┤ ņ×¼ņĪ░ļ¬ģļÉśĻ│Ā ņ׳ņ¢┤, ņØ┤ ļČäņĢ╝ņŚÉ ļīĆĒĢ£ ļ│┤ļŗż Ļ╣ŖņØĆ ņØ┤ĒĢ┤ļź╝ ĒåĄĒĢ┤ ļæÉĻ▓ĮļČĆņĢöņØś ņ╣śļŻīņŚÉ ļīĆĒĢ£ ņāłļĪ£ņÜ┤ ņĀæĻĘ╝ņØä ĒĢ┤ ļ│╝ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż. ļ│Ė ņóģņäżņŚÉņä£ļŖö ĒŖ╣Ē׳ ņ¦Ćļ░®ņé░ņØś ĒĢ®ņä▒, ņé░ĒÖö, ĻĘĖļ”¼Ļ│Ā ņĀĆņןņŚÉ ņ┤łņĀÉņØä ļ¦×ņČöņ¢┤ ņĢöņäĖĒż ļé┤ņŚÉņä£ ļ│┤ņØ┤ļŖö ļīĆņé¼ņŚÉ ļīĆĒĢ┤ ņé┤ĒÄ┤ļ│┤Ļ│Ā, ņØ┤ļź╝ ĒåĄĒĢ┤ ņĢöņØś ņ”ØņŗØĻ│╝ ņ¦äĒ¢ēņØä ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ļŖö ņĀäļץļōżņØä ļ¬©ņāēĒĢ┤ļ│┤Ļ│Āņ×É ĒĢ£ļŗż.
ļ│Ė ļĪĀ
ņĢöņäĖĒżņØś ĒŖ╣ņØ┤ņĀü ļŗ╣ļīĆņé¼(Characteristic glucose metabolism in cancer cells)
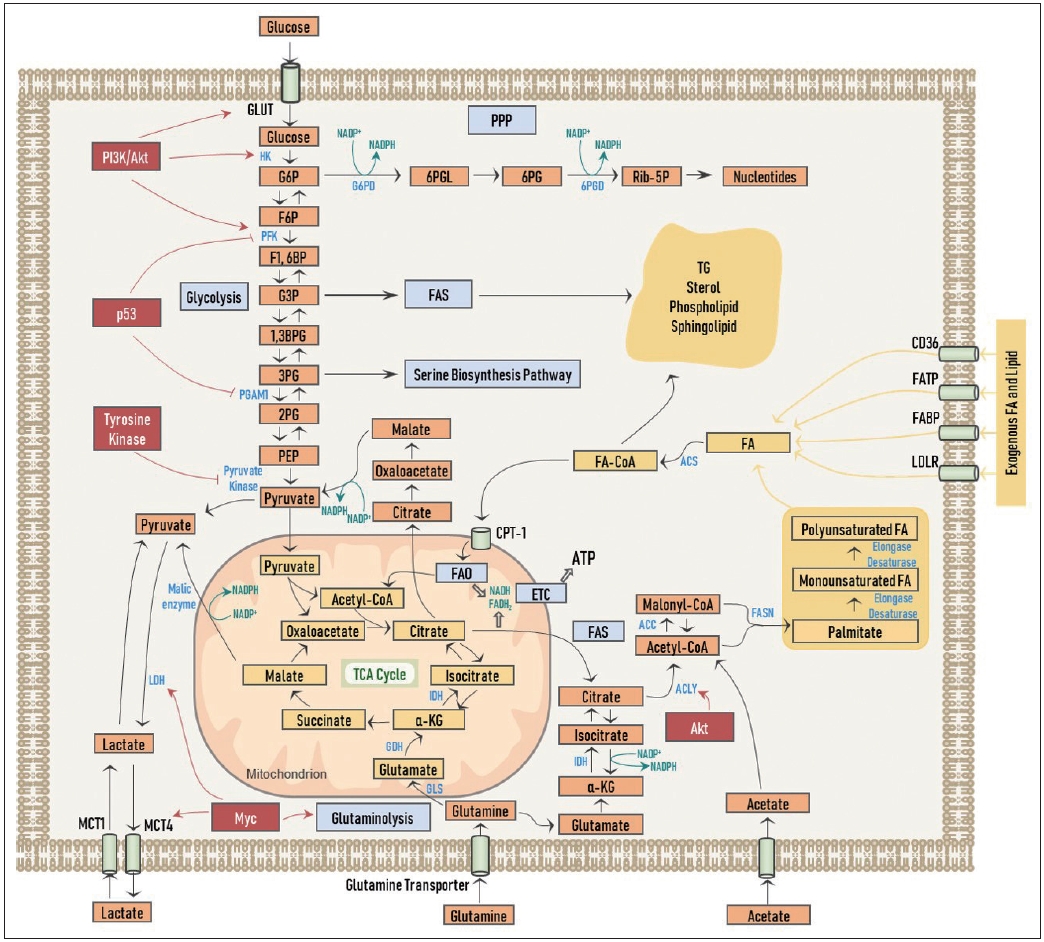
ņĢöņäĖĒżļŖö ņ”ØņŗØĻ│╝ ņä▒ņןņØä ņ£äĒĢ┤ņä£ ļŗ╣ļīĆņé¼(glucose metabolism)ļź╝ ĒÖ£ņÜ®ĒĢśĻ▓ī ļÉśļŖöļŹ░, ņØ┤ļŖö ņŚ¼ļ¤¼ ņĢö ļÅīņŚ░ļ│ĆņØ┤ņŚÉ ļö░ļźĖ ņŗĀĒśĖ ņĀäļŗ¼ņØś Ļ░ĢĒÖöļź╝ ĒåĄĒĢ┤ ņØ╝ņ¢┤ļéśĻ▓ī ļÉ£ļŗż. ņØ┤ļ¤¼ĒĢ£ ĒśäņāüņØä ņ×äņāüņĀüņ£╝ļĪ£ ņĀüņÜ®ĒĢ£ ņśłĻ░Ć ņĢöņäĖĒżĻ░Ć ĒżļÅäļŗ╣ņØś ņ£Āņé¼ņ▓┤ņØĖ 18F-fluorodeoxyglucose ļź╝ ĒØĪņłśĒĢ©ņ£╝ļĪ£ņŹ©, ņĢöņØś ņ¦äļŗ© ļ░Å ņ╣śļŻī ņĀäĒøä ļ░śņØæņä▒ ĒÅēĻ░Ć ļō▒ņŚÉ ĒÖ£ņÜ®ĒĢĀ ņłś ņ׳ļŖö Ļ▓Ćņé¼ņØĖ ņ¢æņĀäņ×Éļ░®ņČ£ļŗ©ņĖĄņ┤¼ņśüņłĀ(positron emission tomography, PET)ņØ┤ļŗż. ņŚ¼ĒĢśĒŖ╝ ņĢöņäĖĒżņØś ņŚ¼ļ¤¼ ĻĄ¼ņä▒ ņÜöņåī(building block)ņØś ĒĢ®ņä▒ņØ┤ ĒĢäņÜöĒĢ£ ņāüĒÖ®ņŚÉņä£, ņäĖĒż ļé┤ļĪ£ ĒØĪņłśļÉ£ ĒżļÅäļŗ╣ņØ┤ ņśżļĪ£ņ¦Ć ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ņØś OXPHOS Ļ│╝ņĀĢņØä ĒåĄĒĢ┤ ļČäĒĢ┤ļÉśņ¢┤ ļ¦ÄņØĆ ņ¢æņØś ATPļź╝ ņāØņé░ĒĢśļŖö Ļ▓ā ļ¦īņ£╝ļĪ£ļŖö ļÅäņøĆņØ┤ ļÉśņ¦Ć ņĢŖĻ│Ā, ĒżļÅäļŗ╣ņØś Ļ│ĀļČäņ×É ņĀäĻĄ¼ļ¼╝ņ¦łļĪ£ņØś ņĀäĒÖśņØ┤ ĒĢäņłśņĀüņØĖļŹ░, Ļ░ĆļĀ╣ ņ¦Ćļ░®ņé░ ĒĢ®ņä▒ņØä ņ£äĒĢ£ acetyl-CoA, ļ╣äĒĢäņłś ņĢäļ»ĖļģĖņé░ ĒĢ®ņä▒ņØä ņ£äĒĢ£ ĒĢ┤ļŗ╣ ņżæĻ░äņ▓┤(glycolytic intermediates), ĻĘĖļ”¼Ļ│Ā ļē┤Ēü┤ļĀłņśżĒŗ░ļō£ņØś ĒĢ®ņä▒ņØä ņ£äĒĢ£ ļØ╝ņØ┤ļ│┤ņśżņŖż(ribose)ņØś ņāØņä▒ņØ┤ ĻĘĖĻ▓āņØ┤ļŗż(Fig. 1).
Ļ░üņóģ ņä▒ņן ņØĖņ×É ļ░Å PI3K/Akt ļō▒ņØś ņóģņ¢æ ņŗĀĒśĖ ņĀäļŗ¼ņŚÉ ņØśĒĢ┤ ĒżļÅäļŗ╣ ņłśņåĪņ▓┤Ļ░Ć ĒÖ£ņä▒ĒÖöļÉśņ¢┤ ĒżļÅäļŗ╣ ņäĖĒż ļé┤ļĪ£ņØś ĒØĪņłśĻ░Ć ņ”ØĻ░ĆļÉśĻ│Ā, ĒŚźņåīĒéżļéśņĢäņĀ£(hexokinase), ĒżņŖżĒżĒöäļŻ®ĒåĀĒéżļéśņĢäņĀ£(phosphofructokinase, PFK)ļź╝ ļ╣äļĪ»ĒĢ£ ĒĢ┤ļŗ╣ ņ×æņÜ®ņØś ņŻ╝ņÜö ĒÜ©ņåīļōżņØś ĻĖ░ļŖź ņĪ░ņĀłņØä ĒåĄĒĢ┤ ĒĢ┤ļŗ╣ ņ×æņÜ®ņØĆ Ļ░ĢĒÖöļÉ£ļŗż[9,10]. ļŹö ļéśņĢäĻ░Ć, ĒāĆņØ┤ļĪ£ņŗĀņØĖņé░ĒÖöĒÜ©ņåī(tyrosine kinase) ņä▒ņ¦łņØä Ļ░Ćņ¦ĆļŖö ļ¦ÄņØĆ ņłśņØś ņĢöņ£ĀņĀäņ×ÉļōżņØĆ Ēö╝ļŻ©ļĖīņé░ņŚ╝ ĒéżļéśņĢäņĀ£(pyruvate kinase)ņØś ĒÖ£ņä▒ņØä ĒŖ╣ņØ┤ņĀüņ£╝ļĪ£ ņ¢ĄņĀ£ĒĢśņŚ¼ ņØ┤ļ¤¼ĒĢ£ ņØ╝ļĀ©ņØś ĒśäņāüļōżņØ┤ ļŗ╣ņżæĻ░äļīĆņé¼ņ▓┤ņØĖ ĒżļÅäļŗ╣-6-ņØĖņé░(glucose-6-phosphate, G6P)ņØ┤ ĒÄ£ĒåĀņśżņŖż ņØĖņé░ Ļ▓ĮļĪ£(pentose phosphate shunt, PPP)ļĪ£ņØś ņ£Āņ×ģņØä ļŖśļĀż ļē┤Ēü┤ļĀłņśżĒŗ░ļō£ ĒĢ®ņä▒ņØä ņÜ®ņØ┤ĒĢśĻ▓ī ĒĢ£ļŗż(Fig. 1). ņØ┤ Ļ│╝ņĀĢņŚÉņä£ ĒżļÅäļŗ╣-6-ņØĖņé░ĒāłņłśņåīĒÜ©ņåī(G6P dehydrogenase, G6PD)ņÖĆ 6-ĒżņŖżĒżĻĖĆļŻ©ņĮśņé░ĒāłņłśņåīĒÜ©ņåī(6-phosphogluconate dehydrogenase, 6PGD) ļæÉ ĒÜ©ņåīņŚÉ ņØśĒĢ┤ nicotinamide adenine dinucleotide phosphate(NADPH) ļśÉĒĢ£ ņāØņä▒ļÉśļŖöļŹ░, ņØ┤ļź╝ ĒåĄĒĢ┤ņä£ ĒĢ®ņä▒ņŚÉ ĒĢäņÜöĒĢ£ ņĀäņ×É(electrons)ļź╝ ņĀäļŗ¼ļ░øĻ▓ī ļÉśĻ│Ā, ļśÉĒĢ£ ņé░ĒÖö ĒÖśņøÉ ņĪ░ņĀł(redox control)ņØä ĒĢĀ ņłś ņ׳Ļ▓ī ļÉ£ļŗż. ļ┐Éļ¦ī ņĢäļŗłļØ╝ ĒĢ┤ļŗ╣ ņżæĻ░äņ▓┤ ņżæņØś ĒĢśļéśņØĖ 3-ņØĖĻĖĆļ”¼ņäĖļź┤ņé░(3-phosphoglycerate)ņØś ņāØņä▒ ņŚŁņŗ£ ņ”ØĻ░ĆļÉśņ¢┤ ņäĖļ”░ ņāØĒĢ®ņä▒ Ļ▓ĮļĪ£(serine biosynthesis pathway)ļź╝ ĒåĄĒĢ┤ ļ╣äĒĢäņłś ņĢäļ»ĖļģĖņé░ ņāØņä▒ņØä ļŖśļ”¼Ļ▓ī ļÉ£ļŗż. ņĢöņ¢ĄņĀ£ ņØĖņ×ÉļĪ£ ņל ņĢīļĀżņ¦ä p53ņØĆ PFKņÖĆ ņØĖņé░ĻĖĆļ”¼ņäĖļź┤ņé░ļ│Ćņ£äĒÜ©ņåī-1 (phosphoglycerate mutase-1)ņØä ņ¢ĄņĀ£ĒĢśņŚ¼ ĒĢ┤ļŗ╣ ņżæĻ░äņ▓┤Ļ░Ć Ļ░üĻ░ü PPPņÖĆ ņäĖļ”░ ņāØĒĢ®ņä▒ Ļ▓ĮļĪ£ļĪ£ņØś ņ£Āņ×ģņØä ņ”ØĻ░Ćņŗ£Ēé©ļŗż(Fig. 1). ļśÉĒĢ£ ņ¦Ćļ░®ņé░ ĒĢ®ņä▒ņØä ņ£äĒĢ┤ ĒĢäņÜöĒĢ£ ĒāäņåīļŖö ĒżļÅäļŗ╣ņØś acetyl-CoAļĪ£ņØś ņĀäĒÖśņØä ĒåĄĒĢ┤ņä£ ņ¢╗ņØä ņłś ņ׳Ļ▓ī ļÉ£ļŗż.
ĒĢ┤ļŗ╣ ņ×æņÜ®ņØä ĒåĄĒĢ┤ ņāØņä▒ļÉ£ Ēö╝ļŻ©ļĖīņé░ņŚ╝ņØĆ ņĀ¢ņé░ĒāłņłśņåīĒÜ©ņåī(lactate dehydrogenase, LDH)ļź╝ ĒåĄĒĢ┤ ņĀ¢ņé░(lactate)ņ£╝ļĪ£ ļ│ĆĒÖśļÉśĻ│Ā, ņØ┤ļĪ£ņŹ© Ļ│╝ļŗżĒĢ£ ĒāäņåīļŖö ļ░░ņČ£ļÉśĻ│Ā ļśÉĒĢ£ NADPHņØś ņāØņä▒ņØä ļÅĢļŖöļŗż. ņĀ¢ņé░ņØĆ ļ¬©ļģĖņ╣┤ļ│ĄņŗżļĀłņØ┤ĒŖĖ ņłśņåĪņ▓┤(monocarboxylate transporter, MCT)ņØś ĒĢśļéśņØĖ MCT4ļź╝ ĒåĄĒĢ┤ ņĢöņäĖĒż ļ░¢ņ£╝ļĪ£ ļ░░ņČ£ļÉśņ¦Ćļ¦ī, ņØ╝ļČĆ ņĢöņäĖĒżņŚÉ ļö░ļØ╝ņä£ļŖö Ļ▒░ĻŠĖļĪ£ MCT1ņØä ĒåĄĒĢ┤ ļŗżņŗ£ ņĢöņäĖĒż ļé┤ļĪ£ ĒØĪņłśĒĢśņŚ¼ ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ OXPHOS Ļ│╝ņĀĢņŚÉ ņØ┤ņÜ®ļÉ£ļŗżļŖö ņŚ░ĻĄ¼ļÅä ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż[11]. ņĢöņäĖĒżņŚÉņä£ļŖö Mycļź╝ ĒåĄĒĢ┤ LDHņÖĆ MCTņØś ļ░£ĒśäņØ┤ ņ”ØĻ░ĆļÉśņ¢┤ ņ׳ļŗż[12]. MycļŖö ļśÉĒĢ£ ĻĖĆļŻ©ĒāĆļ»╝(glutamine)ņØś ņäĖĒż ļé┤ ĒØĪņłśņÖĆ ļČäĒĢ┤(glutaminolysis)ļź╝ ĒÖ£ņä▒ĒÖöņŗ£ņ╝£ ĻĖĆļŻ©ĒāĆļ»╝ņé░(glutamate)ņØ┤ ļ¦Éļ”░ņé░ņŚ╝(malate)Ļ│╝ Ēö╝ļŻ©ļĖīņé░ņŚ╝(pyruvate)ņØä Ļ▒░ņ╣śļ®┤ņä£ NADPHņØś ņāØņä▒ņØä ņ┤ēņ¦äņŗ£Ēé©ļŗż(Fig. 1).
ļīĆņé¼ņØś ņ×¼ĒÄĖņä▒(Reprogramming of metabolism)
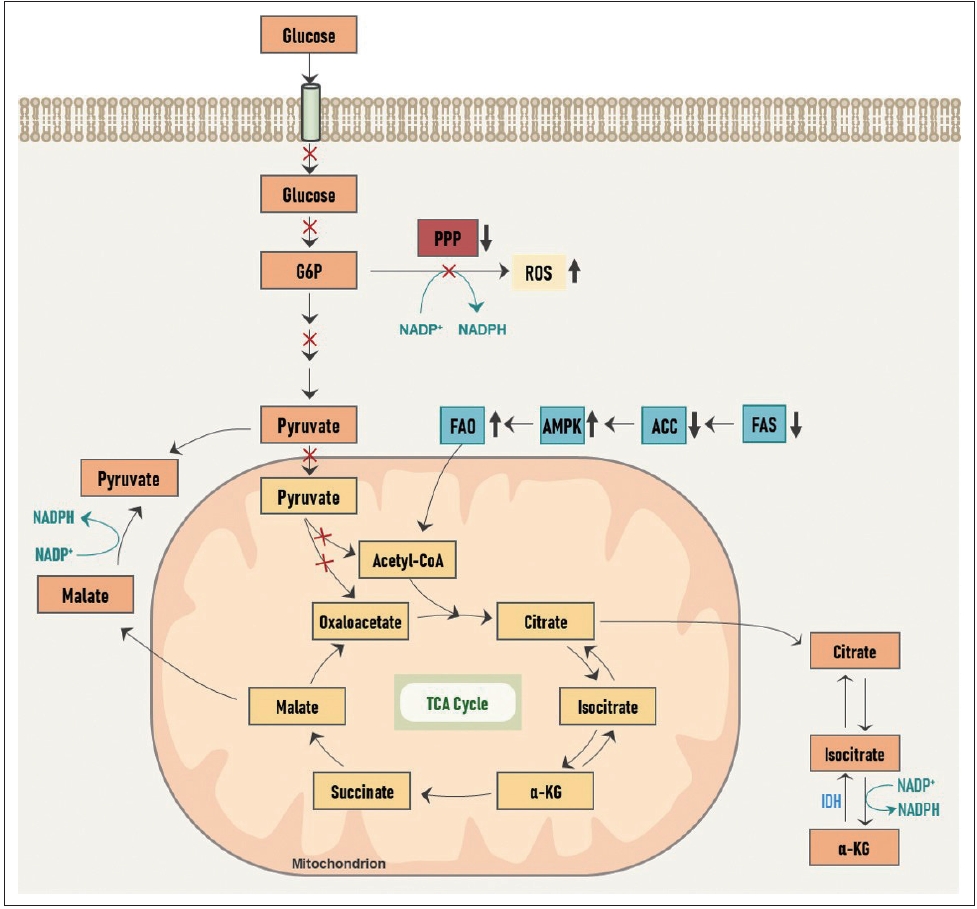
ņä▒ņןĒĢśļŖö ņĢöņäĖĒżņŚÉņä£ ĒĢ┤ļŗ╣ Ļ│╝ņĀĢņØä ņ”ØņŗØņä▒ ļīĆņé¼(proliferative metabolism)ļĪ£ ĒÖ£ņÜ®ĒĢśļŖö Ļ▓āņŚÉ ļ░śĒĢ┤, ņĀäņØ┤ņä▒ ņ╣©ņŖĄņä▒ ņĢöņäĖĒżņŚÉņä£ļŖö ļŗżļźĖ ļ░®Ē¢źņ£╝ļĪ£ ļīĆņé¼Ļ░Ć ņ×¼ĒÄĖņä▒ļÉ£ļŗż(Fig. 2). ņØ┤ļÅÖ ņżæņØĖ ņĀäņØ┤ņĢöņäĖĒżņŚÉņä£ļŖö ĒżļÅäļŗ╣ņØś ĒØĪņłśĻ░Ć Ļ░ÉņåīļÉśĻ│Ā, ņØ┤ļĪ£ ņØĖĒĢ┤ PPPļĪ£ļČĆĒä░ņØś NADPH ņāØņé░ņØ┤ Ļ░ÉņåīļÉśņ¢┤ ņäĖĒż ļé┤ ņé░ĒÖö ĒÖśņøÉ ļŖźļĀźņØ┤ ļ¢©ņ¢┤ņĀĖ ROS ļ░£ņāØņØ┤ ņ”ØĻ░ĆĒĢśĻ▓ī ļÉ£ļŗż. Ļ▓īļŗżĻ░Ć ATP ņāØņé░ ļśÉĒĢ£ Ļ░ÉņåīĒĢśĻ▓ī ļÉśņ¢┤ ņØ┤ļ¤¼ĒĢ£ ņØ╝ļĀ©ņØś ņŚÉļäłņ¦Ć, ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖż(energetic and oxidative stress)ļĪ£ ņØĖĒĢ┤ ņĢöņäĖĒżļŖö ņäĖĒżņé¼ņØś ņ£äĻĖ░ņŚÉ ļåōņØ┤Ļ▓ī ļÉ£ļŗż. ĻĘĖļ¤¼ļéś ĻĘĖņŚÉ ļö░ļØ╝ ņĢöņäĖĒż ļé┤ņŚÉņä£ļŖö ATPņØś Ļ░Éņ¦Ćņ×É(sensor)ņØĖ adenosine monophosphate(AMP)-ĒÖ£ņä▒ ļŗ©ļ░▒ņ¦ł ņØĖņé░ĒÖöĒÜ©ņåī(AMP-activated protein kinase, AMPK)Ļ░Ć ĒÖ£ņä▒ĒÖöļÉśĻ│Ā acetyl-CoA ņ╣┤ļ│ĄņŗżĒÖöĒÜ©ņåī(acetyl-CoA carboxylase, ACC)Ļ░Ć ņ¢ĄņĀ£ļÉśņ¢┤ ņ¦Ćļ░®ņé░ ĒĢ®ņä▒(fatty acid synthesis, FAS)ņØĆ ņ¢ĄņĀ£ļÉśĻ│Ā ņ¦Ćļ░®ņé░ ņé░ĒÖö(fatty acid oxidation, FAO)ļŖö ĒÖ£ņä▒ĒÖöļÉ£ļŗż. FASĻ░Ć Ļ░ÉņåīĒĢ©ņ£╝ļĪ£ņŹ© NADPH ņåīļ¬©ļ¤ēņØĆ ņżäņ¢┤ļōżĻ│Ā, FAOĻ░Ć ņ”ØĻ░ĆĒĢ©ņ£╝ļĪ£ņŹ© ļ¦Éļ”░ņé░ņŚ╝ņŚÉņä£ Ēö╝ļŻ©ļĖīņé░ņŚ╝ņ£╝ļĪ£ņØś ņĀäĒÖśĻ│╝ ņĢäņØ┤ņåīĻĄ¼ņŚ░ņé░ņŚ╝(isocitrate)ņŚÉņä£ ņĢīĒīī-ņ╝ĆĒåĀĻĖĆļŻ©ĒāĆļź┤ņé░(╬▒-ketoglutarate, ╬▒-KG)ļĪ£ņØś ņĀäĒÖśņØ┤ Ļ░üĻ░ü ĒÖ£ņä▒ĒÖöļÉśĻ│Ā, ņØ┤ Ļ│╝ņĀĢņŚÉņä£ NADPH ņāØņé░ņØ┤ ņ”ØĻ░ĆĒĢśņŚ¼ ļČĆņĪ▒ĒĢ£ ņ¢æņØä ļ│┤ņČ®ĒĢĀ ņłś ņ׳Ļ▓ī ļÉ£ļŗż. ļśÉĒĢ£ FAOļź╝ ĒåĄĒĢ┤ ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ OXPHOSļź╝ ĒåĄĒĢ┤ ļ¦ÄņØĆ ņ¢æņØś ATPļź╝ ņāØņé░ĒĢśĻ▓ī ļÉ£ļŗż. ņØ┤ņ▓śļ¤╝ ņ╣©ņŖĄņä▒ ļīĆņé¼(invasive metabolism)ņŚÉņä£ļŖö ņ¦Ćļ░®ņé░ņØś ļīĆņé¼Ļ░Ć ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢ£ļŗż.
ņĢöņäĖĒżņŚÉņä£ņØś ņ¦Ćļ░®ļīĆņé¼(Lipid metabolism in cancer cells)
ņ¦Ćņ¦łņØĆ ļŗżņ¢æĒĢ£ ĻĄ░ņ£╝ļĪ£ ņØ┤ļŻ©ņ¢┤ņ¦ä ņåīņłśņä▒ņØś ņāØļČäņ×Éļ¼╝ņ¦łļĪ£ņä£ ņØĖņ¦Ćņ¦ł(phospholipids), ņŖżĒĢæĻ│Āņ¦Ćņ¦ł(sphingolipids), ĒŖĖļ”¼ņĢäņŗżĻĖĆļ”¼ņäĖļĪż(triacylglycerols, TG), ņŖżĒģīļĪż(sterols) ļō▒ņØä ĒżĒĢ©ĒĢśļ®░ ņ¦Ćļ░®ņé░ņØ┤ ĻĖ░ļ│Ė ĒĢ®ņä▒ļŗ©ņ£äĻ░Ć ļÉ£ļŗż. ņ¦Ćļ░®ņé░ņØĆ ļŗżņ¢æĒĢ£ ĒāäņåīņØś ņłśļź╝ Ļ░Ćņ¦ĆļŖö ĒāäĒÖöņłśņåī ņé¼ņŖ¼ņØ┤ ĻĘĖ ļ¦Éļŗ©ņŚÉ ņ╣┤ļ│Ąņŗ£ĻĖ░(carboxyl group)Ļ░Ć ļČÖņ¢┤ ņ׳ļŖö ĻĄ¼ņĪ░ļĪ£ ņØ┤ļŻ©ņ¢┤ņĀĖ ņ׳ņ£╝ļ®░ ļŗżņ¢æĒĢ£ ĒżĒÖöļÅäļź╝ ļ│┤ņØĖļŗż. ņäĖĒż ļé┤ņŚÉņä£ ņ¦Ćļ░®ņé░ņØĆ ņäĖĒżļ¦ēņØś ĻĄ¼ņä▒ ņä▒ļČäņØ┤ ļÉśĻĖ░ļÅä ĒĢśĻ│Ā, ņŚÉļäłņ¦Ć ļīĆņé¼ ļ░Å ņĀĆņן, ĻĘĖļ”¼Ļ│Ā ņŗĀĒśĖņĀäļŗ¼ļ¼╝ņ¦łļĪ£ņŹ©ļÅä ĒÖ£ņÜ®ņØ┤ ļÉ£ļŗż. ņĢöņäĖĒżņŚÉņä£ņØś ņ¦Ćļ░®ņé░ ļīĆņé¼ ņżæ ļīĆļČĆļČä ņŚ░ĻĄ¼ņØś ņ┤łņĀÉņØĆ ņŻ╝ļĪ£ FASņŚÉ ļ¦×ņČöņ¢┤ņĪīņ£╝ļéś, ņĄ£ĻĘ╝ ļōżņ¢┤ ņĢöņŚÉņä£ņØś FAOņØś ņŚŁĒĢĀņŚÉ ļīĆĒĢ┤ņä£ļÅä Ļ┤Ćņŗ¼ņØ┤ ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ļŗż.
ņ¦Ćļ░®ņé░ ĒĢ®ņä▒(FAS)
ņĢöņäĖĒżĻ░Ć ņ¦Ćļ░®ņé░ņØä ņ¢╗ļŖö ļ░®ļ▓ĢņØĆ ņäĖĒż ņÖĖļČĆļĪ£ļČĆĒä░ ĒØĪņłśļź╝ ĒĢśĻ▒░ļéś, ņäĖĒż ļé┤ņŚÉņä£ ņ¦üņĀæ ĒĢ®ņä▒ņØä ĒĢśļŖö 2Ļ░Ćņ¦Ć Ļ▓ĮļĪ£ļĪ£ņŹ© Ļ░ĆļŖźĒĢśļŗż(Fig. 1). ņĀĢņāü ņäĖĒżņŚÉņä£ļŖö ņŻ╝ļĪ£ ņ¦Ćļ░®ņé░ņØś ņÖĖņØĖņä▒ ĒØĪņłśļź╝ ĒåĄĒĢ┤ ņ¦Ćļ░® ĒĢäņÜöļ¤ēņØä ļ│┤ņČ®ĒĢśļŖö ļŹ░ ļ░śĒĢ┤ņä£ ņĢöņäĖĒżņŚÉņä£ļŖö ņāüļīĆņĀüņ£╝ļĪ£ ņ¦Ćļ░®ņé░ņØś ņŗĀņāØĒĢ®ņä▒(de novo FAS)ņØ┤ ĒÖ£ņä▒ĒÖöļÉśņ¢┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ņŚÉļŖö ņĢöņäĖĒżņŚÉņä£ļÅä ņŻ╝ļ│Ć ĒÖśĻ▓ĮņØś ņ¦Ćļ░®ņé░ņØä ĒĢäņÜöņŚÉ ļö░ļØ╝ ĒÖ£ļ░£Ē׳ ĒØĪņłśĒĢśĻ│Ā, ņĢöņØś ņ¦äĒ¢ēņŚÉ ņ׳ņ¢┤ ņØ┤ļ¤¼ĒĢ£ ņÖĖņØĖņä▒ ņ¦Ćļ░®ņé░ ĒØĪņłśņÖĆ ņ¦Ćļ░®ņé░ņØś ņŗĀņāØĒĢ®ņä▒ ļ¬©ļæÉĻ░Ć ņżæņÜöĒĢśļŗżļŖö ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ļōżņØ┤ ļŖśĻ│Ā ņ׳ļŗż[13,14]. ĒŖ╣Ē׳, ņĢöņäĖĒżļŖö ĒĢäņÜöņŚÉ ļö░ļØ╝ ņŻ╝ņ£ä ņóģņ¢æļ»ĖņäĖĒÖśĻ▓ĮņØś ņ¦Ćļ░®ņäĖĒż(adipocyte)ļĪ£ļČĆĒä░ ņ£Āļ”¼ļÉ£ ņ¦Ćļ░®ņé░ņØä ĒØĪņłśĒĢśņŚ¼ ņ¦Ćļ░®ļīĆņé¼ņŚÉ ĒÖ£ņÜ®ĒĢĀ ņłś ņ׳ļŗż[15-18]. ņ¦Ćļ░®ņé░ņØä ņäĖĒż ļé┤ļĪ£ ĒØĪņłśĒĢśļŖö ņŚŁĒĢĀņØä ļŗ┤ļŗ╣ĒĢśļŖö ļīĆĒæ£ņĀüņØĖ ņłśņåĪņ▓┤ļĪ£ļŖö CD36, ņ¦Ćļ░®ņé░ņłśņåĪļŗ©ļ░▒ņ¦ł(fatty acid transport proteins, FATPs), ņ¦Ćļ░®ņé░Ļ▓░ĒĢ®ļŗ©ļ░▒ņ¦ł(fatty acid binding proteins, FABPs), low-density lipoprotein receptor ļō▒ņØ┤ ņ׳ļŗż.
FASņØś ņ▓½ ļŗ©Ļ│äļŖö ĻĄ¼ņŚ░ņé░ņŚ╝(citrate)ņ£╝ļĪ£ņä£, ņØ┤ļŖö ņŻ╝ļĪ£ ņĢ×ņä£ ņé┤ĒÄ┤ļ│Ė ĒĢ┤ļŗ╣ Ļ│╝ņĀĢņØś ņé░ļ¼╝ņØĖ Ēö╝ļŻ©ļĖīņé░ņŚ╝ņØ┤ TCA ĒÜīļĪ£ļź╝ Ļ▓Įņ£ĀĒĢśņŚ¼ ņāØņä▒ļÉ£ļŗż. ĻĘĖļ¤¼ļéś ņĀĆņé░ņåī ĒÖśĻ▓ĮņØ┤ļéś ļ»ĖĒåĀņĮśļō£ļ”¼ņĢäņØś ĻĖ░ļŖźņØ┤ ņ¢ĄņĀ£ļÉśņ¢┤ ņ׳ļŖö Ļ▓ĮņÜ░ņŚÉ TCA ĒÜīļĪ£Ļ░Ć ļÅīņĢäĻ░ł ņłś ņŚåĻ▓ī ļÉśņ¢┤, ņĢöņäĖĒżļŖö ļīĆņ▓┤ ņśüņ¢æņåīļĪ£ņä£ ĻĖĆļŻ©ĒāĆļ»╝ņØä ņäĖĒż ļé┤ļĪ£ ĒØĪņłśĒĢśņŚ¼ ╬▒-KGļź╝ ņāØņä▒ĒĢśĻ│Ā, ņĢäņØ┤ņåīĻĄ¼ņŚ░ņé░ņŚ╝ ĒāłņłśņåīĒÜ©ņåī(isocitrate dehydrogenase, IDH)ņŚÉ ņØśĒĢ£ ĒÖśņøÉņä▒ ņ╣┤ļ│Ąņŗ£ĒÖö ļ░śņØæ(reductive carboxylation)ļź╝ ĒåĄĒĢ┤ ĻĄ¼ņŚ░ņé░ņŚ╝ņØä ņāØņä▒ĒĢśĻ▓ī ļÉ£ļŗż[19]. ĻĄ¼ņŚ░ņé░ņŚ╝ņØĆ ņØ┤ņ¢┤ņä£ ATP-ĻĄ¼ņŚ░ņé░ņŚ╝ ļČäĒĢ┤ĒÜ©ņåī(ATP-citrate lyase, ACLY)ņŚÉ ņØśĒĢ┤ acetyl-CoAļĪ£ ņĀäĒÖśļÉśļŖöļŹ░, ņĢöņäĖĒżņŚÉņä£ļŖö Aktļź╝ ĒåĄĒĢ┤ ACLYņØś ĒÖ£ņä▒ĒÖöļź╝ ņ┤ēņ¦äņŗ£ņ╝£ acetyl-CoAņØś ĒĢ®ņä▒ņØä ņ”ØĻ░Ćņŗ£Ēé©ļŗż[20]. ņĢöņäĖĒżļŖö ļĢīļĪ£ļŖö MCTļź╝ ĒåĄĒĢ┤ ņÖĖļČĆņŚÉņä£ ņĢäņäĖĒŖĖņé░ņŚ╝(acetate)ņØä ĒØĪņłśĒĢśņŚ¼ acetyl-CoAļź╝ ņ¦üņĀæ ņāØņä▒ĒĢśĻĖ░ļÅä ĒĢ£ļŗż. ĒĢ┤ļŗ╣ Ļ│╝ņĀĢņØ┤ ņāüļīĆņĀüņ£╝ļĪ£ ņĀĆņĪ░ĒĢśĻ│Ā ņ¦Ćļ░®ņé░ ļīĆņé¼Ļ░Ć ĒÖ£ņä▒ĒÖöļÉśņ¢┤ ņ׳ļŖö ņĢöņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ ĒśäņāüņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņśüņāüĒĢÖņĀüņØĖ ņ¦äļŗ© Ļ▓Ćņé¼Ļ░Ć Ļ░ĆļŖźĒĢ£ļŹ░, C11-acetate PET Ļ▓Ćņé¼Ļ░Ć ĻĘĖ ņśłņØ┤ļŗż[21]. Acetyl-CoAļŖö ņØ┤ņ¢┤ņä£ ņåŹļÅäņĀ£ĒĢ£ĒÜ©ņåī(rate-limiting enzyme)ņØĖ ACCņŚÉ ņØśĒĢ┤ malonyl-CoAļĪ£ ņĀäĒÖśļÉ£ļŗż. Malonyl-CoAļĪ£ļČĆĒä░ ņ¦Ćļ░®ņé░ ĒĢ®ņä▒ĒÜ©ņåī(fatty acid synthase, FASN)ņŚÉ ņØśĒĢ┤ ļ╣äļĪ£ņåī ĒżĒÖöņ¦Ćļ░®ņé░ņØĖ Ēīöļ»ĖĒģīņØ┤ĒŖĖ(palmitate)Ļ░Ć ņāØņä▒ņØ┤ ļÉ£ļŗż. Ēīöļ»ĖĒģīņØ┤ĒŖĖļŖö ņØ┤ĒøäņŚÉ ņŚ¼ļ¤¼ ņŗĀņןĒÜ©ņåī(elongase)ņÖĆ ļČłĒżĒÖöĒÜ©ņåī(desaturase)ņØś ņ×æņÜ®ņŚÉ ņØśĒĢ┤ ļŗżņ¢æĒĢ£ ĒāäņåīņØś ņłśņÖĆ ĒżĒÖöļÅäļź╝ Ļ░Ćņ¦ĆļŖö ņ¦Ćļ░®ņé░ļōżņØä ņāØņä▒ĒĢśĻ▓ī ļÉ£ļŗż. ņØ┤ņÖĆ Ļ░ÖņØ┤ ņāØņä▒ļÉ£ ņ¦Ćļ░®ņé░ļōżņØĆ ņĢäņŗż-CoA ĒĢ®ņä▒ĒÜ©ņåī(acyl-CoA synthetase)ņŚÉ ņØśĒĢ┤ fatty acid-CoAļĪ£ Ļ░ĆĻ│ĄņØ┤ ļÉśņ¢┤ ņäĖĒżļ¦ēņØś ĻĄ¼ņä▒ ņä▒ļČäņ£╝ļĪ£ ņżæņÜöĒĢ£ ņØĖņ¦Ćņ¦łņØ┤ļéś ņŖżĒĢæĻ│Āņ¦Ćņ¦łļĪ£ ņĀäĒÖśļÉśĻ▒░ļéś, TGņÖĆ ņŖżĒģīļĪżļĪ£ ļ░öļĆīņ¢┤ ņ¦Ćļ░® ļ░®ņÜĖ(lipid droplet, LD)ņØś ĒśĢĒā£ļĪ£ ņĀĆņןņØ┤ ļÉ£ļŗż. ņŚÉļäłņ¦Ć ņĀĆņןņåīņØĖ LDļŖö ņČöĒøäņŚÉ ņĢöņäĖĒżĻ░Ć ĒĢäņÜöņŗ£ FAOņØś ņŚ░ļŻīļĪ£ņä£ Ļ│ĄĻĖēĒĢśĻ▓ī ļÉ£ļŗż.
ACLY, ACC, FASN ļō▒ FASņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ņŻ╝ņÜö ĒÜ©ņåīļōżņØĆ ņŖżĒģīļĪż ņĪ░ņĀł ņÜöņåī Ļ▓░ĒĢ® ļŗ©ļ░▒ņ¦ł-1(sterol regulatory element binding protein-1, SREBP-1)ņØ┤ļØ╝ļŖö ņĀäņé¼ņĪ░ņĀłņØĖņ×ÉņŚÉ ņØśĒĢ┤ ļ░£ĒśäļÉśĻ│Ā, SREBPļŖö PI3K/Akt/mTORC ņóģņ¢æ ņŗĀĒśĖņŚÉ ņØśĒĢ┤ ĒÖ£ņä▒ĒÖöļÉ£ļŗż.
ņäĖĒż ļé┤ņŚÉņä£ļŖö ĒżĒÖöņ¦Ćļ░®ņé░ ļīĆļ╣ä ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ļ╣äņ£©ņØ┤ ņØ╝ņĀĢĒĢśĻ▓ī ņ£Āņ¦ĆļÉśļŖö Ļ▓āņØ┤ ņżæņÜöĒĢ£ļŹ░, ĒżĒÖöņ¦Ćļ░®ņé░ņØ┤ Ļ│╝ļÅäĒĢśĻ▓ī ņīōņØ┤Ļ▓ī ļÉśļ®┤ ņØ┤ļĪ£ ņØĖĒĢ┤ ļ»ĖĒåĀņĮśļō£ļ”¼ņĢäņØś ĻĖ░ļŖź ņØ┤ņāü, ROS ņ”ØĻ░Ć, ņåīĒżņ▓┤(endoplasmic reticulum, ER) ņŖżĒŖĖļĀłņŖż ļō▒ņØ┤ ļ░£ņāØĒĢśņŚ¼ ņäĖĒżņŚÉ ņ╣śļ¬ģņĀüņØĖ ņåÉņāüņØä Ļ░ĆņĀĖļŗżņżä ņłś ņ׳Ļ▓ī ļÉ£ļŗż. ļö░ļØ╝ņä£ ņ¦Ćļ░®ņé░ņØś ĒāłĒżĒÖö(desaturation)ļź╝ ņ£ĀļÅäĒĢśļŖö ņŖżĒģīņĢäļĪ£ņØ╝-CoA ļČłĒżĒÖöĒÜ©ņåī(stearoyl-CoA desaturase, SCD)ņØś ņŚŁĒĢĀņØ┤ ņżæņÜöĒĢ£ļŹ░, ņØ┤ ĒÜ©ņåīļŖö ĒŖ╣ņØ┤ņĀüņ£╝ļĪ£ ņé░ņåīĻ░Ć ņČ®ļČäĒĢ£ ĒÖśĻ▓ĮņŚÉņä£ ņ×æņÜ®ņØ┤ ņøÉĒÖ£ĒĢśĻ▓ī ņØ┤ļŻ©ņ¢┤ņ¦äļŗż. ņĢöņäĖĒżņŚÉņä£ļŖö SCDņØś ļ░£ĒśäņØ┤ ņ”ØĻ░ĆļÉśņ¢┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░[22,23], ņĀĆņé░ņåī ĒÖśĻ▓ĮņŚÉ ļåōņØĖ ņĢöņäĖĒżņŚÉņä£ļŖö SCDņØś ĒÖ£ņä▒ņØ┤ ņ¢ĄņĀ£ļÉśņ¢┤ ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ņāØņä▒ņØ┤ Ļ░ÉņåīĒĢśĻĖ░ņŚÉ, ņØ┤ļ¤¼ĒĢ£ ņāüĒÖ®ņŚÉ ļīĆļ╣äĒĢśņŚ¼ ņĢöņäĖĒżļŖö LD ļé┤ņØś ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ņ£Āļ”¼ļź╝ ņ”ØĻ░Ćņŗ£ĒéżĻ▒░ļéś[24] ņĀĆņé░ņåīņ”Øņ£Āļ░£ņØĖņ×É-1╬▒(hypoxiainducible factor-1╬▒) ļ░£Ēśä ņ”ØĻ░Ćļź╝ ĒåĄĒĢ┤ ņÖĖļČĆ ĒÖśĻ▓Įņ£╝ļĪ£ļČĆĒä░ ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ĒØĪņłśļź╝ ļŖśļĀżņä£ ņ¦Ćļ░®ļÅģņä▒(lipotoxicity)ņ£╝ļĪ£ļČĆĒä░ ņäĖĒżļź╝ ļ│┤ĒśĖĒĢśĻ▓ī ļÉ£ļŗż[25].
ĒżĒÖöņ¦Ćļ░®ņé░Ļ│╝ ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ņäĖĒż ņĪ░ņä▒ņØĆ ņäĖĒżļ¦ēņØś ņ£ĀļÅÖņä▒(fluidity)ņŚÉļÅä Ļ╣ŖņØĆ ņśüĒ¢źņØä ļ»Ėņ╣£ļŗż. ņ”ē, ĒżĒÖöņ¦Ćļ░®ņé░ņØś ļ╣äņ£©ņØ┤ ļåÆņØäņłśļĪØ ņäĖĒżļ¦ēņØĆ Ļ▓¼Ļ│ĀĒĢ┤ņ¦ĆĻ│Ā ņ£ĀļÅÖņä▒ņØĆ Ļ░ÉņåīĒĢśĻ▓ī ļÉśĻ│Ā, ņØ┤ļ¤¼ĒĢ£ Ļ▓ĮņÜ░ ņĢöņäĖĒżļŖö ņĢĮļ¼╝ ĒØĪņłś Ļ░ÉņåīļĪ£ ņØĖĒĢ┤ ĒĢŁņĢöņĀ£ļĪ£ļČĆĒä░ ņĀĆĒĢŁņä▒ņØä Ļ░Ćņ¦ł ņłś ņ׳Ļ▓ī ļÉ£ļŗż[26]. ļ░śļ®┤ņŚÉ, ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś ļ╣äņ£©ņØ┤ ļåÆņØĆ ņĢöņäĖĒżņŚÉņä£ļŖö ņäĖĒżļ¦ēņØś ĻĄ¼ņä▒ņØ┤ ļŖÉņŖ©ĒĢ┤ņĀĖ ņ£ĀļÅÖņä▒ņØĆ ņ”ØĻ░ĆĒĢśņŚ¼ ņØ┤ļĪ£ ņØĖĒĢ┤ ņĢöņäĖĒżņØś ņØ┤ļÅÖĻ│╝ ņĀäņØ┤ļź╝ ņÜ®ņØ┤ĒĢśĻ▓ī ĒĢ£ļŗż[27]. ņØ┤ņ▓śļ¤╝, ņ¦Ćļ░®ņé░ņØś ņĪ░ņä▒ņŚÉ ļö░ļźĖ ņäĖĒżļ¦ēņØś ņ£ĀļÅÖņä▒ņØä ņĪ░ņĀłĒĢśņŚ¼ ņĢöņäĖĒżļŖö Ļ░ü ĒĢäņÜö ņāüĒÖ®ņŚÉ ļ¦×ļÅäļĪØ ņØ┤ņĀÉņØä ņĘ©ĒĢĀ ņłś ņ׳Ļ▓ī ļÉ£ļŗż.
ņ¦Ćļ░®ņé░ ņé░ĒÖö(FAO)
ņĢ×ņä£ ņ¢ĖĻĖēĒĢ£ ļ░öņÖĆ Ļ░ÖņØ┤, ņĀäņØ┤ņä▒ ņĢöņäĖĒżņŚÉņä£ļŖö ĒżļÅäļŗ╣ņØś ĒØĪņłśĻ░Ć ņżäņ¢┤ļōżĻ│Ā ņĢöņäĖĒżņØś ņØ┤ļÅÖĻ│╝ ņ╣©ņŖĄņØś Ļ│╝ņĀĢņŚÉņä£ ļ¦ÄņØĆ ņ¢æņØś ATPļź╝ ĒĢäņÜöļĪ£ ĒĢśĻ▓ī ļÉśļŖöļŹ░, ņØ┤ļ¤¼ĒĢ£ ņāüĒÖ®ņŚÉņä£ Ļ░Ćņן ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ņŚÉļäłņ¦Ćļź╝ ņāØņé░ĒĢĀ ņłś ņ׳ļŖö ļ░®ļ▓ĢņØ┤ FAOņØ┤ļŗż. ĒāäņłśĒÖöļ¼╝Ļ│╝ ļ╣äĻĄÉĒĢ┤ņä£ ņ¦Ćļ░®ņé░ņØĆ 2ļ░░ļéś ļ¦ÄņØĆ ņ¢æņØś ATPļź╝ ņĀ£Ļ│ĄĒĢĀ ņłś ņ׳ļŗż. ļæÉĻ▓ĮļČĆņĢöņäĖĒż ņżæņŚÉņä£ļÅä ņĀäņØ┤ļź╝ ņ£Āļ░£ĒĢśļŖö ņØ╝ļČĆ ņäĖĒżļōżņŚÉņä£ ņ¦Ćļ░®ņé░ņØś ņłśņÜ®ņ▓┤ņØĖ CD36ņØś ļ░£ĒśäĻ│╝ ĒĢ©Ļ╗ś ņ¦Ćļ░®ļīĆņé¼ Ļ┤ĆļĀ© ņ£ĀņĀäņ×ÉņØś ļ░£ĒśäņØ┤ ļåÆņØĆ Ļ▓āņØä ĒÖĢņØĖĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņ׳ņ£╝ļ®░[13], ņØ┤ ĒŖ╣ņĀĢ ņĢöņäĖĒżĻĄ░ņŚÉ Ēīöļ»ĖĒģīņØ┤ĒŖĖ Ēś╣ņØĆ Ļ│Āņ¦Ćļ░®ņŗØņØ┤(high fat diet) ņ▓śļ”¼ ņŗ£ņŚÉ ņøÉļ░£ ļČĆņ£ä ņĢöņØś ļ│ĆĒÖöļŖö ļ»Ėļ»ĖĒĢśĻ│Ā, ĒŖ╣ņØ┤ĒĢśĻ▓ī ņĢöņäĖĒżņØś ņĀäņØ┤ļ¦īņØä ņ┤ēņ¦äņŗ£ņ╝░Ļ│Ā, ļ░śļīĆļĪ£ CD36ņØä ņ¢ĄņĀ£ ņŗ£ņŚÉ ņøÉļ░£ ļČĆņ£äņØś ņĢöņŚÉ ļīĆĒĢ£ ņśüĒ¢źņŚåņØ┤ ņĢöņØś ņĀäņØ┤ļ¦ī ņäĀĒāØņĀüņ£╝ļĪ£ ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ņØīņØä ĒÖĢņØĖĒĢ£ ļ░ö ņ׳ļŗż. ļ┐Éļ¦ī ņĢäļŗłļØ╝, ņøÉļ░£ ļČĆņ£äņØś ņĢöĻ│╝ ļ╣äĻĄÉĒĢśņŚ¼ ļ”╝ĒöäņĀłļĪ£ ņĀäņØ┤Ļ░Ć ņØ┤ļŻ©ņ¢┤ņ¦ä ņĢöņäĖĒżļź╝ ļČäņäØĒĢ£ Ļ▓░Ļ│╝, FAOņÖĆ Ļ┤ĆļĀ©ļÉ£ ņ£ĀņĀäņ×ÉņØś ļ░£ĒśäņØ┤ ņ”ØĻ░ĆļÉśņ¢┤ ņ׳ņ£╝ļ®░, ņØ┤ļŖö ĒŖ╣ņØ┤ĒĢśĻ▓ī ĒÖ£ņä▒ĒÖöļÉ£ yesņŚ░Ļ┤Ć ļŗ©ļ░▒ņ¦ł(yes-associated protein, YAP)ņØ┤ļØ╝ļŖö ņĀäņé¼ņØĖņ×ÉņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ©ņØä ĒÖĢņØĖĒĢ£ ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ļÅä ņ׳ļŗż[28]. ņØ┤ ņŚ░ĻĄ¼ņŚÉņä£ļÅä FAOļź╝ ņ¢ĄņĀ£ĒĢśļŖö ņĢĮļ¼╝ņØä Ēł¼ņŚ¼ĒĢśĻ▒░ļéś YAP ļ░£ĒśäņØä ņ¢ĄņĀ£Ē¢łņØä ļĢī ļ”╝ĒöäņĀł ņĀäņØ┤ļź╝ ņäĀĒāØņĀüņ£╝ļĪ£ ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ņŚłļŗż. ņØ┤ļź╝ ĒåĀļīĆļĪ£ ļ│╝ ļĢī FAOļź╝ ņ¢ĄņĀ£ĒĢśļŖö Ļ▓āņØ┤ ņĢöņØś ņ╣©ņŖĄĻ│╝ ņĀäņØ┤ļź╝ ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ļŖö ĒÜ©Ļ│╝ņĀüņØĖ ņĀäļץņ×äņØä ņ¦Éņ×æĒĢ┤ ļ│╝ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż.
ņ¦Ćļ░®ņé░ņØĆ ņØ┤ņ▓śļ¤╝ CD36ņØä ĒåĄĒĢ┤ ņÖĖļČĆņŚÉņä£ ņäĖĒż ļé┤ļĪ£ ĒØĪņłśĻ░Ć ļÉśĻ▒░ļéś LD ļé┤ņŚÉ ņĀĆņןļÉśņ¢┤ ņ׳ļŹś TGļĪ£ļČĆĒä░ ņ£Āļ”¼ļÉśņ¢┤, ņ╣┤ļź┤ļŗłĒŗ┤Ēīöļ»ĖĒåĀņØ╝ ņĀäņØ┤ĒÜ©ņåī(carnitine palmitoyl transferase-1)ņŚÉ ņØśĒĢ┤ ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ļĪ£ ņØ┤ļÅÖĒĢśņŚ¼ FAOĻ░Ć ņØ┤ļŻ©ņ¢┤ņ¦äļŗż. FAO Ēś╣ņØĆ ļ▓ĀĒāĆ ņé░ĒÖöĻ│╝ņĀĢ(╬▓-oxidation)ņØś Ļ▓░Ļ│╝ļĪ£ acetyl-CoA, nicotinamide adenine dinucleotide(NADH), flavin adenine dinucleotide(FADH2)Ļ░Ć ņāØņä▒ļÉśĻ│Ā, acetyl-CoAļŖö TCA ĒÜīļĪ£ļĪ£ ņ£Āņ×ģļÉśņ¢┤ NADHĻ░Ć ņāØņä▒ļÉśĻ│Ā, NADHņÖĆ FADH2ļŖö ņĀäņ×ÉņĀäļŗ¼Ļ│ä(electron transport chain, ETC)ļĪ£ ņ£Āņ×ģļÉśņ¢┤ ATPĻ░Ć ļ░£ņāØĒĢśĻ▓ī ļÉ£ļŗż(Fig. 1). ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļé┤ acetyl-CoAĻ░Ć TCA ĒÜīļĪ£ļź╝ ĒåĄĒĢ┤ NADHļź╝ ņāØņä▒ĒĢśņ¦Ćļ¦ī, ĒĢ©Ļ╗ś ĻĄ¼ņŚ░ņé░ņŚ╝ ļśÉĒĢ£ ņāØņä▒ļÉśņ¢┤ ņäĖĒżņ¦łļĪ£ ņ£ĀņČ£ļÉśĻ│Ā, ņØ┤ņ¢┤ņä£ ļŗżņØīņØś ļæÉ Ļ░Ćņ¦Ć Ļ▓ĮļĪ£ļĪ£ ņŚ░Ļ▓░ļÉśņ¢┤ ĻĘĖ Ļ│╝ņĀĢņŚÉņä£ NADPHĻ░Ć ņāØņä▒ļÉ£ļŗż: 1) Ļ░üĻ░ü ņŚ░ņåŹņĀüņ£╝ļĪ£ ņśźņé┤ļĪ£ņĢäņäĖĒŖĖņé░ņŚ╝(oxaloacetate), ļ¦Éļ”░ņé░ņŚ╝, Ēö╝ļŻ©ļĖīņé░ņŚ╝ņ£╝ļĪ£ ņĀäĒÖśļÉśļŖö Ļ▓ĮļĪ£, 2) IDHņŚÉ ņØśĒĢ┤ ņĢäņØ┤ņåīĻĄ¼ņŚ░ņé░ņŚ╝ņŚÉņä£ ╬▒-KGļĪ£ ļ│ĆĒÖśļÉśļŖö Ļ▓ĮļĪ£. ņØ┤ņ▓śļ¤╝ ņāØņä▒ļÉ£ NADPHļŖö ņĀäņłĀĒĢ£ ļ░öņÖĆ Ļ░ÖņØ┤ ņĀäņØ┤ņä▒ ņĢöņäĖĒżņŚÉņä£ ROS ļ░£ņāØ ņ”ØĻ░ĆļĪ£ ņØĖĒĢ£ ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżļź╝ Ļ░Éņåīņŗ£ĒéżļŖö ĒĢŁņé░ĒÖöņĀ£ļĪ£ņä£ņØś ņŚŁĒĢĀņØä ĒĢśņ¦Ćļ¦ī, ĒĢ£ĒÄĖņ£╝ļĪ£ļŖö ļÅÖĒÖö ņ×æņÜ®ņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ņŚ¼ļ¤¼ ĒÜ©ņåīļōżņŚÉ ļīĆĒĢ£ ņĪ░ĒÜ©ņåīļĪ£ņä£ ņ×æņÜ®ĒĢśņŚ¼ ņ”ØņŗØņä▒ ņĢöņäĖĒżņØś ĒĢ®ņä▒ Ļ│╝ņĀĢņŚÉļÅä ņØ┤ņÜ®ļÉĀ ņłś ņ׳ļŗż. ņÜöņĢĮĒĢśņ×Éļ®┤, ņĢöņäĖĒżņŚÉņä£ļŖö ĒĢäņÜöņŗ£ FAOņØś Ļ│╝ņĀĢņØä ĒåĄĒĢ┤ ATPņÖĆ NADPHļź╝ Ļ│ĄĻĖēļ░øņØä ņłś ņ׳ļŖö Ļ▓āņØ┤ļŗż[29]. ļśÉĒĢ£, FAOļź╝ ĒåĄĒĢ┤ ņāØņä▒ļÉ£ acetylCoAļŖö ĻĄ¼ņŚ░ņé░ņŚ╝ņ£╝ļĪ£ņØś ļ│ĆĒÖś Ļ│╝ņĀĢņØä Ļ▒░ņ│É ņäĖĒżņ¦ł ļé┤ acetylCoAļĪ£ ļŗżņŗ£ ļ│ĆĒÖśļÉśņ¢┤ FASļĪ£ ņØ┤ņ¢┤ņ¦ł ņłśļÅä ņ׳ļŗżļŖö ņĀÉņØä ļ»ĖļŻ©ņ¢┤ ļ│┤ņĢä, FAOņÖĆ FASļŖö ĒĢäņÜöņŚÉ ļö░ļØ╝ ņāüĒśĖ ļ│┤ņÖäņĀüņØĖ Ļ┤ĆĻ│äļĪ£ ņäĖĒżņŚÉ ņ£ĀņÜ®ĒĢśĻ▓ī ņ×æņÜ®ĒĢĀ ņłś ņ׳ļŗż.
ņ¦Ćļ░® ļīĆņé¼ņØś ņ╣śļŻī Ēæ£ņĀü(Therapeutic targets in lipid metabolism)
ņ¦ĆĻĖłĻ╣īņ¦Ć ņé┤ĒÄ┤ļ┤żļō»ņØ┤ ņĢöņäĖĒż ļé┤ņŚÉņä£ļŖö ĒŖ╣ņØ┤ņĀüņØĖ ņ¦Ćļ░® ļīĆņé¼ļź╝ ĒåĄĒĢ┤ ņÖĖļČĆ ĒÖśĻ▓ĮņŚÉ ļö░ļØ╝ ļīĆņé¼ņĀü Ļ░Ćņåīņä▒(metabolic plasticity)ņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņ£╝ļ®░, ļö░ļØ╝ņä£ ņĢöņØś ņØ┤ļ¤¼ĒĢ£ ņāØņĪ┤ ņĀäļץņØä ņŚŁņØ┤ņÜ®ĒĢśņŚ¼ ņĢöņ╣śļŻīņØś Ēæ£ņĀüņ£╝ļĪ£ ļ¬©ņāēĒĢ┤ļ│╝ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż(Table 1).
ņ¦Ćļ░®ņé░ ĒĢ®ņä▒ņØś Ēæ£ņĀüĒÖö(Targeting FAS)
ņÖĖņØĖņä▒ ņ¦Ćļ░®ņé░ņØś ņäĖĒż ļé┤ ĒØĪņłśļź╝ ļŗ┤ļŗ╣ĒĢśļŖö ļ¦ēļŗ©ļ░▒ņ¦łņØä ņ╣śļŻī Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ņĀäļץņØä ĒåĄĒĢ┤ ņĢöņäĖĒżņØś ņ¦Ćļ░®ņé░ Ļ│ĄĻĖēņØä ļ¦ēņØä ņłś ņ׳ļŗż. ĒĢŁCD36 ĒĢŁņ▓┤ļź╝ ĒåĄĒĢ┤ ĒĢŁņĢö ĒÜ©Ļ│╝Ļ░Ć ņ׳ņØīņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░[13,14], FATP ļ░Å FABPņŚÉ ļīĆĒĢ£ ņ¢ĄņĀ£ ņĢĮļ¼╝ņØä ĒåĄĒĢ┤ņä£ļÅä ņĢöņØś ņĀäņØ┤ļź╝ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ņżäņØ╝ ņłś ņ׳ņØīņØä ĒÖĢņØĖĒĢ£ ļ░ö ņ׳ļŗż[15,30].
ņØ┤ņÖĆ ļīĆļ│äĒĢśņŚ¼ ņĢöņäĖĒż ļé┤ņØś ņ¦Ćļ░®ņé░ ņŗĀņāØ ĒĢ®ņä▒ņØä Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ņ╣śļŻī ņĀäļץņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļōżļÅä ĒÖ£ļ░£Ē׳ ņ¦äĒ¢ē ņżæņØ┤ļŗż. ņĀĢņāü ņäĖĒżņŚÉ ļ╣äĒĢ┤ ņĢöņäĖĒż ļé┤ņŚÉņä£ ņ¦Ćļ░®ņé░ņØś ņŗĀņāØ ĒĢ®ņä▒ņØ┤ ņāüļīĆņĀüņ£╝ļĪ£ ĒÖ£ņä▒ĒÖöļÉśņ¢┤ ņ׳ĻĖ░ņŚÉ ļ¦żļĀźņĀüņØĖ ĒĢŁņĢöņĀäļץņ£╝ļĪ£ņä£ ņŻ╝ļ¬®ņØä ļ¦ÄņØ┤ ļ░øĻ│Ā ņ׳ļŗż. ņ¦Ćļ░®ņé░ ņŗĀņāØ ĒĢ®ņä▒ Ļ│╝ņĀĢņŚÉņä£ ņ×æņÜ®ĒĢśļŖö ĒÜ©ņåī ņżæņŚÉņä£ FASNņØä ņ¢ĄņĀ£ĒĢśļŖö ņŚ¼ļ¤¼ ņĢĮļ¼╝ņØ┤ Ļ░£ļ░£ļÉśņ¢┤ ļ¦ÄņØĆ ņŚ░ĻĄ¼Ļ░Ć ņ׳ņŚłļŖöļŹ░, orlistat, cerulenin, C75, IPI-9119, EGCGĻ░Ć ĻĘĖ ņśłņØ┤ļŗż[31-35]. ĻĘĖļ¤¼ļéś ļÅÖļ¼╝ ņŗżĒŚśņØä ĒåĄĒĢ┤ ņØ┤ļ¤¼ĒĢ£ ņĢĮņØä ņé¼ņÜ®Ē¢łņØä ļĢī ĻĖēĻ▓®ĒĢ£ ņ▓┤ņżæ ņåīņŗżņØ┤ļéś ņŗĀĻ▓ĮņżäĻĖ░ņäĖĒżņØś ĻĖ░ļŖź ņØ┤ņāüņ£╝ļĪ£ ņØ╝ļČĆ ņŗ¼Ļ░üĒĢ£ ļČĆņ×æņÜ®ņØ┤ ĒÖĢņØĖļÉśņ¢┤ ņ×äņāü ņĀüņÜ®ņŚÉļŖö ņĢäņ¦ü ņäĖņŗ¼ĒĢ£ ņŻ╝ņØśĻ░Ć ĒĢäņÜöĒĢ£ ņāüĒÖ®ņØ┤ļŗż[32,36]. ļ┐Éļ¦ī ņĢäļŗłļØ╝, FAS ņ¢ĄņĀ£ļź╝ ĒåĄĒĢ┤ ņāüļīĆņĀüņ£╝ļĪ£ FAO ĒÖ£ņä▒ĒÖöļź╝ ĒåĄĒĢ£ ņĢö ņĀäņØ┤Ļ░Ć ņ”ØĻ░ĆļÉśņŚłļŗżļŖö Ļ▓░Ļ│╝ļÅä ņ׳ņ¢┤[37,38], Ļ┤æļ▓öņ£äĒĢ£ ņĢöņØś ņĀüņÜ®ļ│┤ļŗżļŖö ņĢöņØś Ļ░ü ņ¦äĒ¢ē ļŗ©Ļ│äņŚÉ ļ¦×ļŖö ņ╣śļŻī ņĀäļץņØä ņŗ£ņé¼ĒĢ£ļŗż. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ņŚÉļŖö FAS ņ¢ĄņĀ£ļź╝ ĒĢśņ¦Ćļ¦ī Ļ░äņĀæņĀüņØĖ FAO ĒÖ£ņä▒ĒÖö ĒÜ©Ļ│╝ļź╝ ļ│┤ņØ┤ņ¦Ć ņĢŖļŖö TVB-3166ņØ┤ļéś TVB2640Ļ│╝ Ļ░ÖņØĆ ņ░©ņäĖļīĆ FASN ņ¢ĄņĀ£ņĀ£Ļ░Ć Ļ░£ļ░£ļÉśņ¢┤, ņ¦Ćļ░® ņĪ░ņ¦ü ņåīņŗż ļ░Å ņŗ¼Ļ░üĒĢ£ ņ▓┤ņżæ Ļ░Éņåī ļō▒ņØś ļČĆņ×æņÜ® ņŚåņØ┤ ļ│┤ļŗż ĒŖ╣ņØ┤ņĀüņØĖ FAS ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļź╝ ļ│┤ņŚ¼ ĒāüņøöĒĢ£ ĒĢŁņĢöĒÜ©Ļ│╝ņØś Ļ░ĆļŖźņä▒ņØä ļ│┤ņØĖ Ļ▓░Ļ│╝ļÅä ņ׳ļŗż[39,40]. FASN ņØ┤ ņÖĖņŚÉļÅä ACLYņÖĆ ACCļź╝ Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ņ╣śļŻī ņĀäļץņŚÉ ļīĆĒĢ┤ņä£ļÅä ļ¦ÄņØĆ ņŚ░ĻĄ¼Ļ░Ć ļÉśņ¢┤, ņ×äņāü ņĀüņÜ®ņØś ņ×Āņ×¼ņĀü Ļ░ĆļŖźņä▒ņØä ņŚ┐ļ│╝ ņłś ņ׳ļŗż[41-44]. ĒŖ╣Ē׳ļéś epidermal growth factor receptor ņ¢ĄņĀ£ņĀ£ņØĖ cetuximabņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ņØä ļ│┤ņØ┤ļŖö ļæÉĻ▓ĮļČĆņĢöņäĖĒżņŚÉņä£ cetuximabĻ│╝ ĒĢ©Ļ╗ś ACC ņ¢ĄņĀ£ņĀ£ņØĖ TOFAļź╝ ļ│æņÜ®ņ▓śļ”¼ĒĢśņśĆņØä ļĢīņŚÉ ĒÜ©Ļ│╝Ļ░Ć ņ׳ņŚłļŗżļŖö ļ│┤Ļ│ĀļÅä ņ׳ļŗż[45]. ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņĢöņäĖĒżļŖö ņāüĒÖ®ņŚÉ ļö░ļØ╝ ļīĆņ▓┤ņśüņ¢æņåīļĪ£ņä£ ĻĖĆļŻ©ĒāĆļ»╝ņØä ĒØĪņłśĒĢśņŚ¼ ĻĖĆļŻ©ĒāĆļ»╝ļīĆņé¼ Ļ│╝ņĀĢņØä ĒåĄĒĢ┤ TCA ĒÜīļĪ£ļĪ£ ņ£Āņ×ģļÉśĻ▒░ļéś ĻĄ¼ņŚ░ņé░ņŚ╝ņ£╝ļĪ£ ņĀäĒÖśļÉśņ¢┤ FASļĪ£ ņØ┤ņ¢┤ņ¦ĆļŖöļŹ░, ĻĖĆļŻ©ĒāĆļ»╝ ļČäĒĢ┤ ĒÜ©ņåī(glutaminase)ļź╝ ņ¢ĄņĀ£ĒĢśļŖö ņĢĮļ¼╝ņØĖ BPTES ņ▓śļ”¼ļź╝ ĒåĄĒĢ┤ ļæÉĻ▓ĮļČĆņĢöņäĖĒżņØś ņä▒ņןņØä ņĀĆĒĢ┤ĒĢĀ ņłś ņ׳ņŚłļŗżļŖö Ļ▓░Ļ│╝ļÅä ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż[46].
ACLY, ACC, FASN, SCD ļō▒ FA ĒĢ®ņä▒ Ļ│╝ņĀĢņŚÉ Ļ┤ĆļĀ©ļÉ£ ĒÜ©ņåīļōżņØś ļ░£ĒśäņØĆ ņĀäņé¼ņĪ░ņĀłņØĖņ×ÉņØĖ SREBP-1ņŚÉ ņØśĒĢ┤ ņĪ░ņĀłļÉ£ļŗż. ĻĘĖļ¤¼ļéś SREBP-1ļŖö ņĀäņé¼ņØĖņ×ÉņØ┤ĻĖ░ņŚÉ ņØ┤ļź╝ ņ¦üņĀæ ņ¢ĄņĀ£ĒĢśļŖö ņĢĮļ¼╝ Ļ░£ļ░£ņØĆ ņ¢┤ļĀĄĻ│Ā SREBP-1Ļ│╝ Ļ┤ĆļĀ©ļÉ£ ļŗ©Ļ│äņŚÉņä£ ņ£ĀņÜ®ĒĢ£ ņ╣śļŻī ņĀäļץņØä ņāØĻ░üĒĢ┤ļ│╝ ņłś ņ׳ļŗż. SREBP-1ņØ┤ ĒÖ£ņä▒ĒÖöļÉśĻĖ░ ņ£äĒĢ┤ņä£ļŖö SREBP-1ņØ┤ SREBP-ļČäņŚ┤ ĒÖ£ņä▒ĒÖö ļŗ©ļ░▒ņ¦ł(SREBP-cleavage activating protein, SCAP)Ļ│╝ Ļ▓░ĒĢ®ĒĢśņŚ¼ ERņŚÉņä£ Ļ│©ņ¦Ćņ▓┤(golgi apparatus)ļĪ£ ņØ┤ļÅÖņØä ĒĢśĻ▓ī ļÉ£ļŗż. SREBP-1Ļ│╝ SCAPņØś Ļ▓░ĒĢ®ņØä ļ░®ĒĢ┤ĒĢśļŖö ņĢĮļ¼╝ņØ┤ Ļ░£ļ░£ļÉśņ¢┤ ĒĢŁņĢöĒÜ©Ļ│╝Ļ░Ć ņ׳ņØīņØä ĒÖĢņØĖĒĢ£ ļ░ö ņ׳ļŖöļŹ░ fatostatin, betulin ļō▒ņØ┤ ĻĘĖ ņśłņØ┤ļŗż[47-49]. ĻĘĖ ļ░¢ņŚÉļÅä SREBP-1ņØś ņāüņ£ä ņĪ░ņĀłņØĖņ×ÉņØĖ Ļ░ä X ņłśņÜ®ņ▓┤(liver X receptor)ļź╝ ņ¢ĄņĀ£ĒĢśļŖö ņĀäļץņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļÅä ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż[50,51].
ņÜöņ╗©ļīĆ ņĢöņäĖĒżļŖö ņ¦Ćļ░®ņé░ ņŗĀņāØ ĒĢ®ņä▒ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņÖĖņØĖņä▒ ņ¦Ćļ░®ņé░ ĒØĪņłśļź╝ ĒåĄĒĢ┤ņä£ļÅä ņäĖĒżĻ░Ć ĒĢäņÜöĒĢ£ ņ¦Ćļ░®ņé░ Ļ│ĄĻĖēņØä ļ░øņØä ņłś ņ׳ĻĖ░ņŚÉ, ņØ┤ ļæÉ Ļ│╝ņĀĢņØä ļ¬©ļæÉ ļ¦ēļŖö Ļ▓āņØ┤ ļŹö ĒÜ©Ļ│╝ņĀüņØĖ ņ╣śļŻī ņĀäļץņØ╝ ņłś ņ׳ļŗż. ĒĢ£ĒÄĖ, ņĀäņłĀĒĢ£ ļ░öņÖĆ Ļ░ÖņØ┤ ņ¦Ćļ░®ņé░ ĒĢ®ņä▒ņØä ĒåĄĒĢ┤ ņāØņä▒ļÉ£ Ēīöļ»ĖĒģīņØ┤ĒŖĖļŖö ņŚ¼ļ¤¼ ņŗĀņןĒÜ©ņåīņÖĆ ļČłĒżĒÖöĒÜ©ņåīņØś ņ×æņÜ®ņØä ĒåĄĒĢ┤ ļŗżņ¢æĒĢ£ ļŗ©ņØ╝ļČłĒżĒÖöņ¦Ćļ░®ņé░(monounsaturated fatty acid)Ļ│╝ ļŗżļČłĒżĒÖöņ¦Ćļ░®ņé░(polyunsaturated fatty acid)ņØä ņāØņä▒ĒĢśņŚ¼ ņäĖĒżĻ░Ć ĒĢäņÜöĒĢ£ ļŗżņ¢æĒĢ£ ņÜ®ļÅäļĪ£ ĒÖ£ņÜ®ļÉśļŖöļŹ░, ņĢöņäĖĒżņØś ņŗĀņןĒÜ©ņåīņÖĆ ļČłĒżĒÖöĒÜ©ņåīļź╝ Ļ░üĻ░ü ņ¢ĄņĀ£ĒĢśņŚ¼ ņĢöņØś ņ”ØņŗØĻ│╝ ņä▒ņןņØä ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ļŗżļŖö Ļ▓░Ļ│╝Ļ░Ć ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż[52-54].ĒŖ╣Ē׳ ļīĆĒæ£ņĀüņØĖ ļČłĒżĒÖö ĒÜ©ņåīņØĖ SCDņŚÉ ļīĆĒĢ£ ņ¢ĄņĀ£ņĀ£Ļ░Ć ļ¦ÄņØ┤ Ļ░£ļ░£ļÉśĻ│Ā ņŚ░ĻĄ¼ļÉśņŚłļŖöļŹ░, betulinic acid, SSI-4, MF-438 ļō▒ņØ┤ ņ׳ļŗż[55-57]. ĻĘĖļ¤¼ļéś ņØ╝ļČĆ ņĢöņäĖĒżņŚÉņä£ļŖö SCD ņ¢ĄņĀ£ņĀ£ļź╝ ņ▓śļ”¼ĒĢśņśĆņØä ļĢī ņÖĖņØĖņä▒ ņ¦Ćļ░®ņé░ ĒØĪņłśļéś ļŗżļźĖ ļČłĒżĒÖöĒÜ©ņåīņØś ņ×æņÜ®ņØä ĒåĄĒĢ┤ ļČłĒżĒÖöņ¦Ćļ░®ņé░ņØś Ļ│ĄĻĖēņØä ņ£Āņ¦ĆĒĢśņŚ¼ SCD ņ¢ĄņĀ£ņĀ£ņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ņØä ļ│┤ņØĖ Ļ▓ĮņÜ░ļÅä ņ׳ļŗż[25,58].
ņ¦Ćļ░®ņé░ ņé░ĒÖöņØś Ēæ£ņĀüĒÖö(Targeting FAO)
FAOņØś Ļ░Ćņן ņ▓½ ļŗ©Ļ│äņØ┤ņ×É ņåŹļÅä ņĪ░ņĀł ļŗ©Ļ│ä(rate-limiting step)ņØĖ CPT-1ņØś ņ×æņÜ®ņØä ņ¢ĄņĀ£ĒĢśļŖö ņĢĮņĀ£Ļ░Ć Ļ░£ļ░£ļÉśņ¢┤ ņĢöņØś ņä▒ņן ļ░Å ņ¦äĒ¢ēņØä ņĀ£ņ¢┤ĒĢĀ ņłś ņ׳ļŗżļŖö Ļ▓░Ļ│╝ļōżņØ┤ ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŖöļŹ░, etomoxirņÖĆ perhexiline ļō▒ņØ┤ ĻĘĖ ļīĆĒæ£ņĀüņØĖ ņśłņØ┤ļŗż[28,59,60]. ĻĘĖ ļ░¢ņŚÉļÅä FAOņØś ļ¦łņ¦Ćļ¦ē ļŗ©Ļ│äņŚÉņä£ ņ×æņÜ®ĒĢśļŖö ĒÜ©ņåīņØĖ 3-ņ╝ĆĒåĀņĢäņŗżĒŗ░ņś¼ļČäĒĢ┤ĒÜ©ņåī(3-ketoacylthiolase)ņŚÉ ļīĆĒĢ£ ņ¢ĄņĀ£ņĀ£ļÅä Ļ░£ļ░£ļÉ£ ļ░ö ņ׳ļŖöļŹ░ trimetazidine, ranolazine ļō▒ņØ┤ ņ׳ļŗż[61,62]. ļśÉĒĢ£, FAO ņ×Éņ▓┤ļź╝ ņ¢ĄņĀ£ĒĢśĻĖ░ļ│┤ļŗżļŖö ĻĘĖ ņØ┤ĒøäņŚÉ ETCņŚÉņä£ ATPĻ░Ć ņāØņä▒ļÉśļŖö Ļ│╝ņĀĢņØä Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ETC ņ¢ĄņĀ£ņĀ£ļōżņŚÉ ļīĆĒĢ£ ĒĢŁņĢöĒÜ©Ļ│╝ļÅä ļ¦ÄņØ┤ ļ│┤Ļ│ĀļÉśņŚłļŖöļŹ░, ņØĄĒ׳ ļŗ╣ļć©ļ│æ ņ╣śļŻīņĢĮņĀ£ļĪ£ļÅä ņל ņĢīļĀżņ¦ä metforminĻ│╝ phenformin ļō▒ņØ┤ ņ׳ļŗż. MetforminņØĆ ĒŖ╣Ē׳ļéś AMPKļź╝ ĒÖ£ņä▒ĒÖöņŗ£ĒéżļŖö ņ×æņÜ®ļÅä ņ׳ņ¢┤, ņØ┤ļź╝ ĒåĄĒĢ┤ ACCĻ░Ć ņ¢ĄņĀ£ļÉśĻ│Ā ņØ┤ņ¢┤ņä£ FASĻ░Ć ņ░©ļŗ©ļÉśņ¢┤ ĒĢŁņóģņ¢æ ĒÜ©Ļ│╝ļź╝ ļ│┤ņØĖļŗżļŖö Ļ▓░Ļ│╝ļÅä ņ׳ļŗż[63].
ņÜöņĢĮ ļ░Å Ļ▓░ļĪĀ
ņĢöņäĖĒżļŖö ņé░ņåīĻ░Ć ņČ®ļČäĒĢ£ ĒÖśĻ▓ĮņŚÉņä£ļÅä ĒĢ┤ļŗ╣ Ļ│╝ņĀĢĻ│╝ OXPHOSļź╝ ĒåĄĒĢ£ Ēö╝ļŻ©ļĖīņé░ņŚ╝ņØś ņé░ĒÖö Ļ│╝ņĀĢņØä ļČäļ”¼ĒĢ©ņ£╝ļĪ£ņŹ©, ĒāäņłśĒÖöļ¼╝ņØ┤ ņĄ£ļīĆņ╣śņØś ATPļź╝ ņāØņé░ĒĢśļŖö ļīĆņŗĀ ņĢöņäĖĒż ņä▒ņןņŚÉ ĒĢäņÜöĒĢ£ ļÅÖĒÖöņ×æņÜ®ņŚÉ ļŗ╣ļīĆņé¼ļź╝ ĒŖ╣ņØ┤ņĀüņ£╝ļĪ£ ņØ┤ņÜ®ĒĢ£ļŗż. ņØ┤ņÖĆ ļŹöļČłņ¢┤ ņ”ØņŗØņä▒ ņĢöļīĆņé¼(proliferative cancer metabolism)ļŖö ĒÖ£ņä▒ĒÖöļÉ£ FASņØä ĒåĄĒĢ┤ ņŚÉļäłņ¦Ć ņĀĆņן, ņäĖĒżļ¦ē ĒĢ®ņä▒, ņŗĀĒśĖņĀäļŗ¼ ļō▒ņØä ņ£äĒĢ┤ ņ¦Ćļ░®ņé░ņØä ņØ┤ņÜ®ĒĢ£ļŗż. ĻĘĖļ¤¼ļéś ļĢīļĪ£ļŖö ņ¦äĒ¢ēņä▒ ņĢöņŚÉņä£ļŖö ņĢöņäĖĒżņØś ņĀäņØ┤ņÖĆ ņØ┤ļÅÖņØä ņ£äĒĢ┤ ņ╣©ņŖĄņä▒ ļīĆņé¼ļź╝ ĒÖ£ņÜ®ĒĢśĻ▓ī ļÉśļŖöļŹ░, ņ¦Ćļ░®ņé░ņØś ļīĆņé¼Ļ░Ć ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢ£ļŗż. FAOļź╝ ĒåĄĒĢ┤ ļīĆņé¼ņĀü ņŖżĒŖĖļĀłņŖżņŚÉ ļåōņØĖ ņĢöņäĖĒżņŚÉ ATPņÖĆ NADPH Ļ│ĄĻĖēņØä ĒĢĀ ņłś ņ׳Ļ▓ī ļÉ£ļŗż. ņØ┤ņ▓śļ¤╝ ņĢö ņä▒ņןņØś Ļ░ü ļŗ©Ļ│ä, Ēś╣ņØĆ ņĢöņØ┤ ļåōņŚ¼ņ¦ä ĒÖśĻ▓ĮņŚÉ ļ¦×ņČöņ¢┤ FASņÖĆ FAOņØś Ļ░ü ļŗ©Ļ│äļź╝ Ēæ£ņĀüņ£╝ļĪ£ ĒĢ£ ņ╣śļŻīļź╝ Ļ│ĀļĀżĒĢ┤ļ│╝ ņłś ņ׳ņØä Ļ▓āņØ┤Ļ│Ā, ļŗżņ¢æĒĢ£ ņĢöņóģņŚÉ ņĀüņÜ®ĒĢ£ ņŚ¼ļ¤¼ Ļ▓░Ļ│╝Ļ░Ć ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż. ļæÉĻ▓ĮļČĆņĢöņØĆ ņĢäņ¦ü ņāüļīĆņĀüņ£╝ļĪ£ ņĀüņÜ® ļ▓öņ£äĻ░Ć ļäōņ¦Ć ņĢŖņ£╝ļéś, ņØ┤ļ»Ė ĒāĆņĢöņóģņŚÉņä£ ņĀüņÜ®ĒĢ£ ĻĖŹņĀĢņĀüņØĖ Ļ▓░Ļ│╝ļōżņØä ĒåĀļīĆļĪ£ ņ¦Ćļ░®ņé░ ļīĆņé¼ņØś ņāłļĪ£ņÜ┤ ņ╣śļŻī ņĀäļץ Ļ░ĆļŖźņä▒ņØä ĻĖ░ļīĆĒĢ┤ ļ│╝ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż.